Epistasis refers to the non-additivity of fitness effects arising from specific combinations of mutations. (1) outline three types of epistasis: magnitude epistasis (“same direction as expected but are not perfectly additive”), sign epistasis (“effect of one of the substitutions changes direction in the context of the other”), and reciprocal epistasis (“effects of both substitutions change direction when they are made together”).
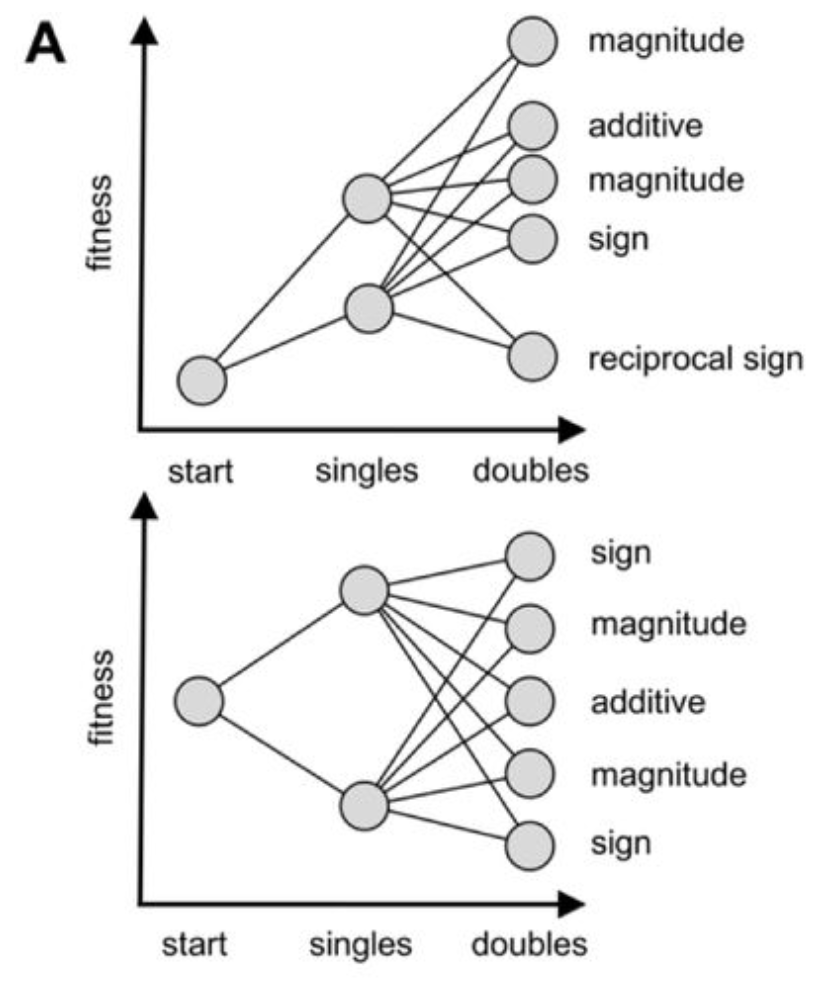 Figure from (1)
Figure from (1)
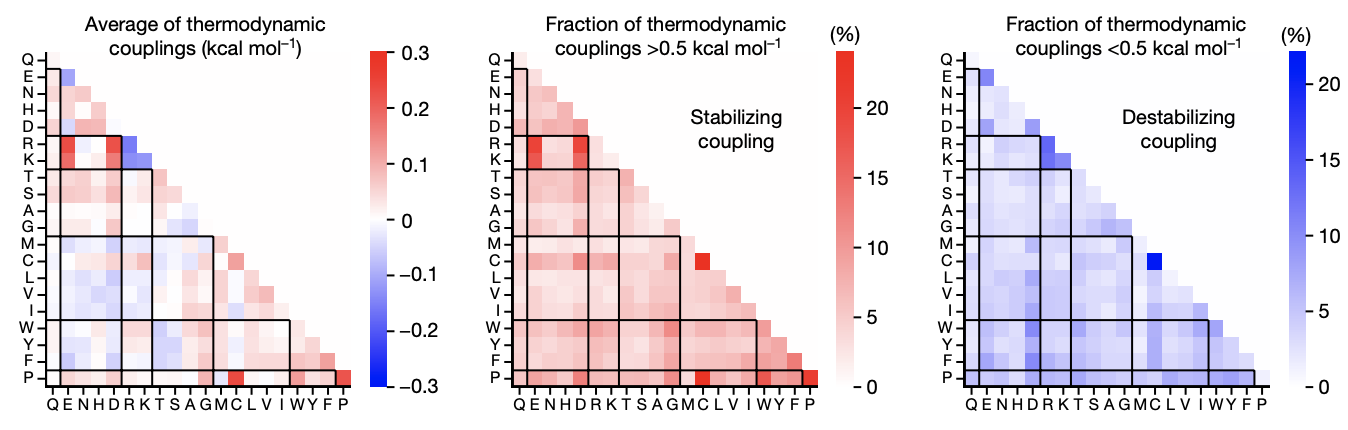 Figure from (2)
Figure from (2)
Notes
- Negative epistasis is approximately 100x more common than positive epistasis (3).
- Rates of magnitude, sign, and reciprocal epistasis are constant across positions (1). These differed from a null additive-only model with noise, which had 74% magnitude, 22% sign, and 4% reciprocal sign epistasis.
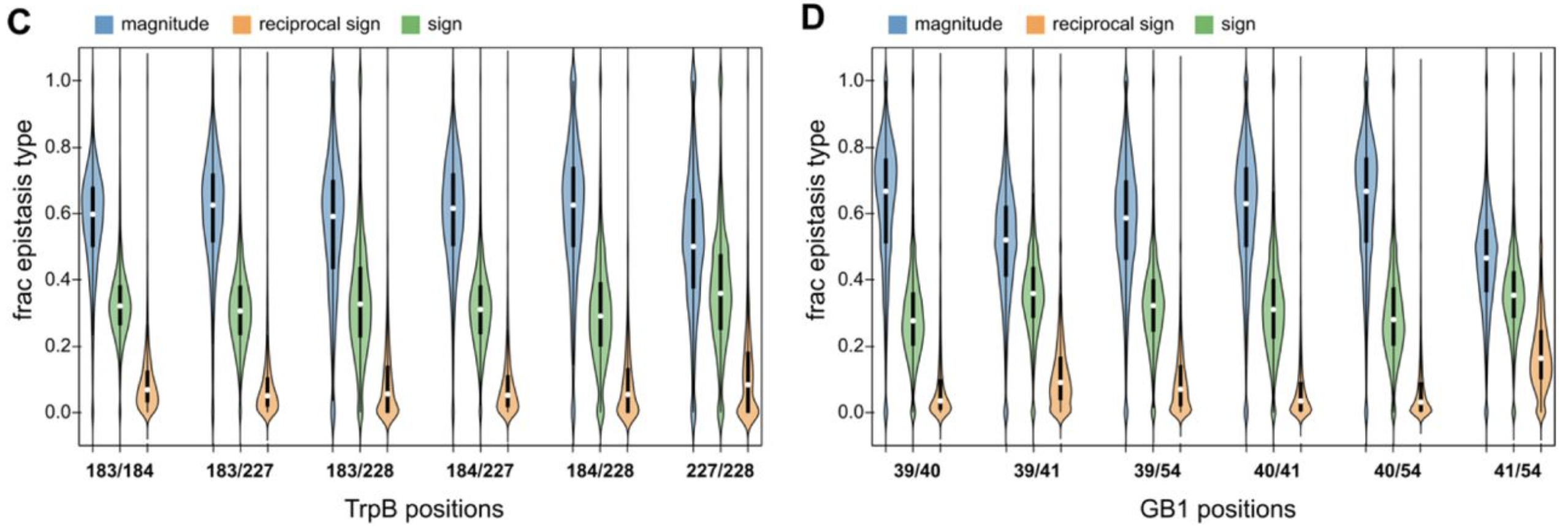 Figure from (1)
Figure from (1)
Examples
- The S373P mutation in the Spike protein is disadvantageous in pre-Omicron-variant versions of SARS-CoV-2 but advantageous in Omicron. Mentioned by Bloom and Neher (4).
- The combination of N501Y and Q498R in the Spike protein of SARS-CoV-2 increases the binding affinity to ACE2 by 387-fold. This is believed to have led to the immune-evasive mutations in the Omicron-variant and was observed in vitro by Zahradník et al. (5) prior to the emergence of Omicron:
- One example of third-order epistasis is the J-domain, where strong non-additivity is observed among a triad, but disappears if any of the three are mutated to alanine (2).
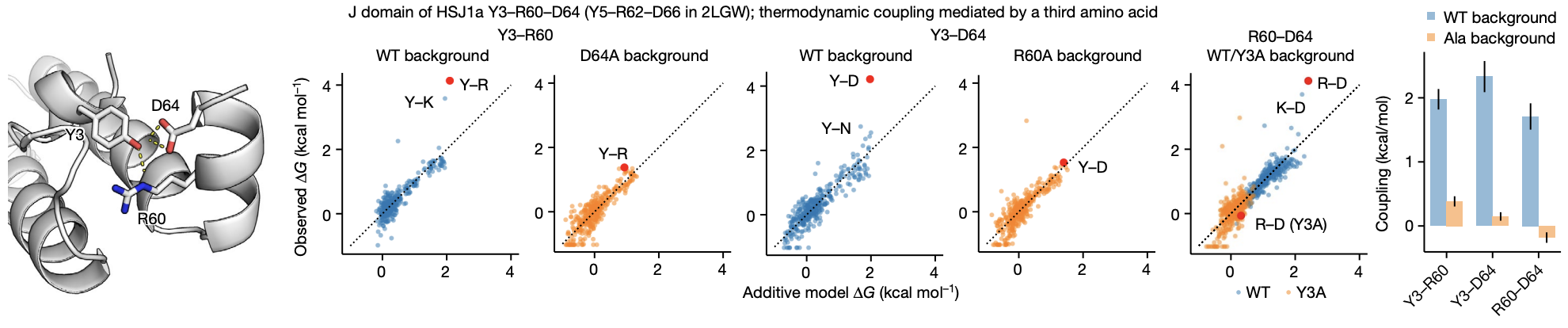 Figure from (2)
Figure from (2)
Measuring epistasis
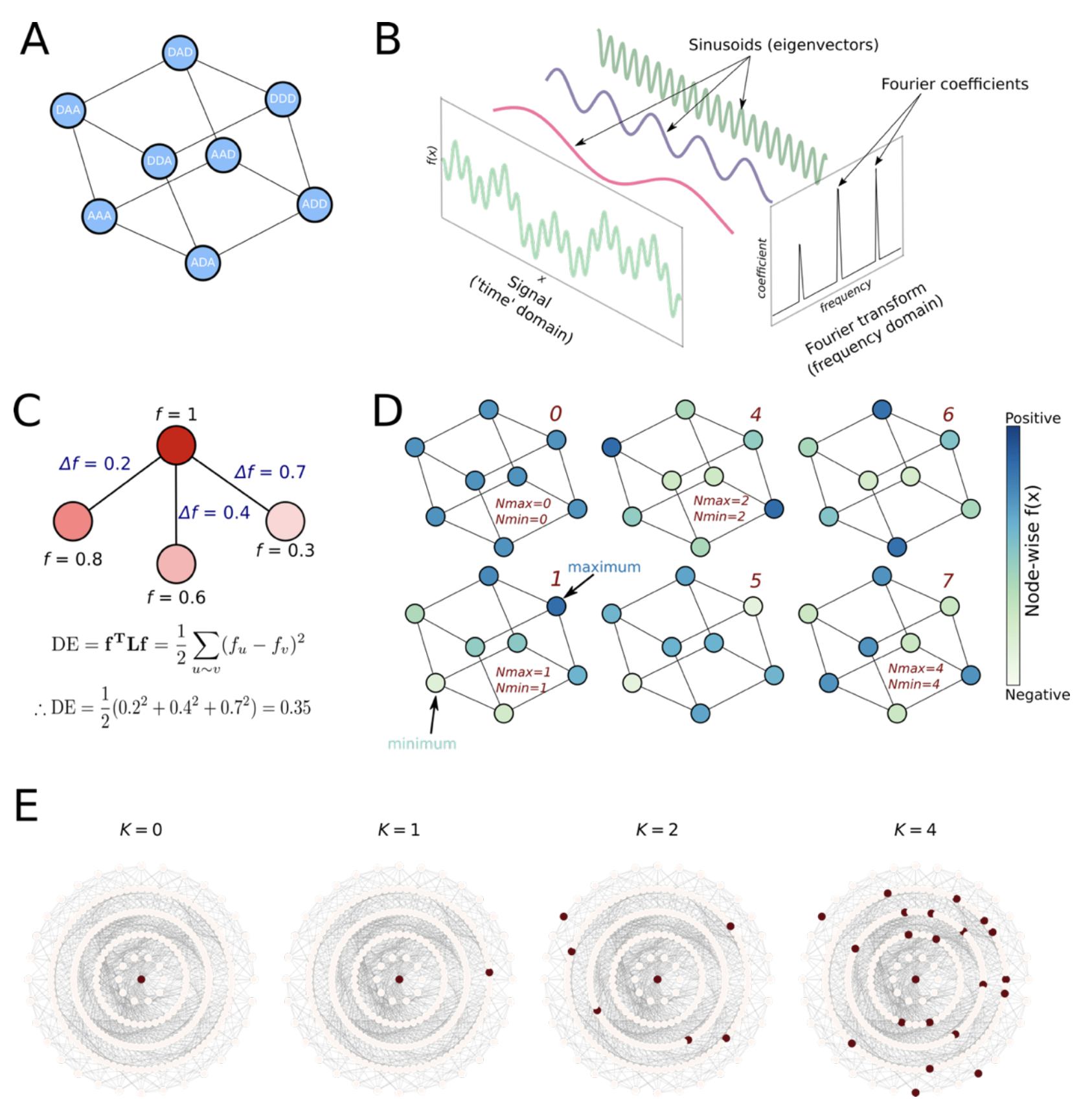 Figure from (6)
Figure from (6)
- Graph Fourier Transform: A decomposition that breaks down the signal (here, fitness) into distinct “epistatic orders”
- Dirichlet energy: quantifies how variable (i.e., rugged) the landscape is, with high energy corresponding to high ruggedness
- NK model: A linear model where is the number of sites and is the number of interacting sites; when , effect of all mutations is linear, whereas larger indicates more interactions and therefore more epistasis
1.
Johnston KE, Almhjell PJ, Watkins-Dulaney EJ, Liu G, Porter NJ, Yang J, et al. A combinatorially complete epistatic fitness landscape in an enzyme active site. Proceedings of the National Academy of Sciences. 2024;121(32). Available from: https://doi.org/10.1073/pnas.2400439121
2.
Tsuboyama K, Dauparas J, Chen J, Laine E, Mohseni Behbahani Y, Weinstein JJ, et al. Mega-scale experimental analysis of protein folding stability in biology and design. Nature. 2023;620(7973):434–44. Available from: https://doi.org/10.1038/s41586-023-06328-6
3.
Sarkisyan KS, Bolotin DA, Meer MV, Usmanova DR, Mishin AS, Sharonov GV, et al. Local fitness landscape of the green fluorescent protein. Nature. 2016;533(7603):397–401. Available from: https://doi.org/10.1038/nature17995
4.
Bloom JD, Neher RA. Fitness effects of mutations to SARS-CoV-2 proteins. Virus Evolution. 2023;9(2):vead055. Available from: https://doi.org/10.1093/ve/vead055
5.
Zahradník J, Marciano S, Shemesh M, Zoler E, Harari D, Chiaravalli J, et al. SARS-CoV-2 variant prediction and antiviral drug design are enabled by RBD in vitro evolution. Nature Microbiology. 2021;6(9):1188–98. Available from: https://doi.org/10.1038/s41564-021-00954-4
6.
Sandhu M, Chen JZ, Matthews DS, Spence MA, Pulsford SB, Gall B, et al. Computational and Experimental Exploration of Protein Fitness Landscapes: Navigating Smooth and Rugged Terrains. Biochemistry. 2025;64(8):1673–84. Available from: https://doi.org/10.1021/acs.biochem.4c00673