The complementarity-determining regions of antibodies, abbreviated CDRs and sometimes collectively called hypervariable regions, are loops that mediate binding to antigens. They are the most difficult part of the protein to predict and get fine-tuned by affinity maturation and somatic hypermutation.
General observations
Confomational clustering
Conformational clusters of CDRs were first described by Chothia & Lesk (1)
CDRH1
- Camelid nanobodies account for most of the diversity in CDRH1 lengths, which in humans and mice are almost all length 7 (2).
CDRH2
- The residue identity at heavy chain residue 71 (Chothia numbering) influences the conformation of CDRH2 (citation needed).
- In Kabat numbering CDRH2 is slightly too short (reason unclear).
- HDX-MS data suggest that CDRH2 has decreased dynamics following Affinity maturation (citation needed).
CDRH3
Conformation and dynamics
CDRL1
- Longer CDRL1 loops correlate with restricted CDRH3 dynamics (3).
CDRL2
- HDX-MS data suggest that CDRH2 has decreased dynamics following affinity maturation (citation needed).
- In Kabat numbering this is slightly too short (reason unclear; citation needed).
CDRL3
- Shorter CDRL3 loops correlate with restricted CDRH3 dynamics (3).
- More variable than CDRL1 or CDRL2 due to VJ recombination (2).
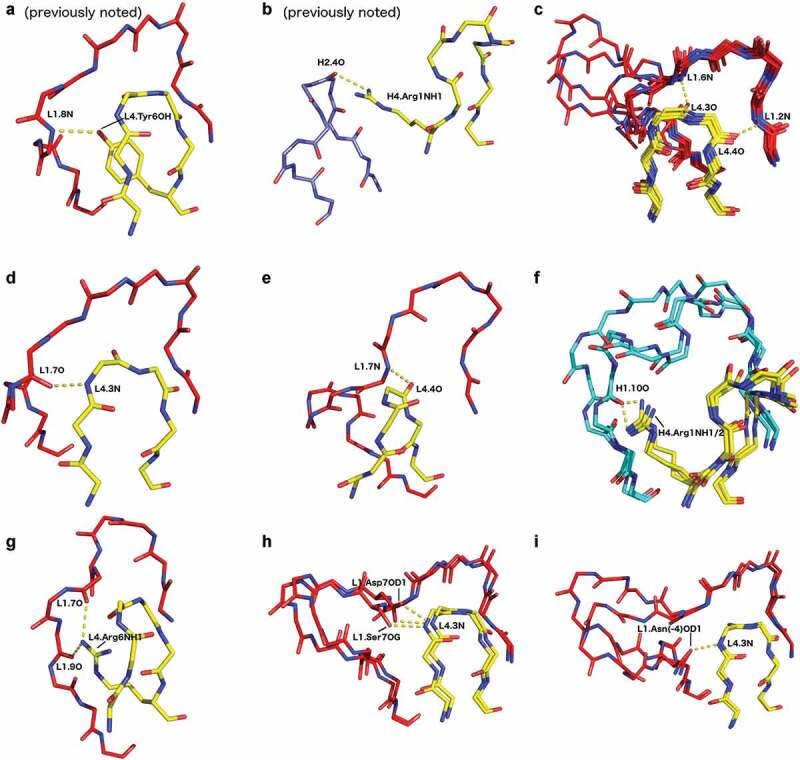
CDR4
Also known as the DE loop
- Sequence variation in the DE loop exceeds that of the framework
- Length variation found in broadly neutralizing antibodies
- Also known as the DE motif, since it connects the D and E strands of Antibodies; the name CDR4 is less common.
- PyIgClassify2 found four distinct clusters for length-6 light chain. Two of these with exclusively kappa subtype, one exclusively with lambda subtype, and one mixed with both; almost all heavy chain structures have length-8 CDR4 and co-cluster (4,5).
- Also used in Nanobodies and can be engineered to improve binding (6)
- Fernandez-Quintero et al. (7) and Kelow et al. (4) found that this can modulate the conformation of other CDRs, particularly CDRL1
1.
Chothia C, Lesk AM. Canonical structures for the hypervariable regions of immunoglobulins. Journal of Molecular Biology. 1987;196(4):901–17. Available from: https://doi.org/10.1016/0022-2836(87)90412-8
2.
Nowak J, Baker T, Georges G, Kelm S, Klostermann S, Shi J, et al. Length-independent structural similarities enrich the antibody CDR canonical class model. mAbs. 2016;8(4):751–60. Available from: https://doi.org/10.1080/19420862.2016.1158370
3.
Guloglu B, Deane CM. Specific attributes of the VL domain influence both the structure and structural variability of CDR-H3 through steric effects. Frontiers in Immunology. 2023;14. Available from: https://doi.org/10.3389/fimmu.2023.1223802
4.
Kelow SP, Adolf-Bryfogle J, Dunbrack RL. Hiding in plain sight: structure and sequence analysis reveals the importance of the antibody DE loop for antibody-antigen binding. mAbs. 2020;12(1). Available from: https://doi.org/10.1080/19420862.2020.1840005
5.
Kelow S, Faezov B, Xu Q, Parker M, Adolf-Bryfogle J, Dunbrack RL. A penultimate classification of canonical antibody CDR conformations. openRxiv; 2022. Available from: https://doi.org/10.1101/2022.10.12.511988
6.
Ketaren NE, Fridy PC, Malashkevich V, Sanyal T, Brillantes M, Thompson MK, et al. Unique Binding and Stabilization Mechanisms Employed By and Engineered Into Nanobodies. openRxiv; 2023. Available from: https://doi.org/10.1101/2023.10.22.563475
7.
Fernández-Quintero ML, Pomarici ND, Math BA, Kroell KB, Waibl F, Bujotzek A, et al. Antibodies exhibit multiple paratope states influencing VH–VL domain orientations. Communications Biology. 2020;3(1). Available from: https://doi.org/10.1038/s42003-020-01319-z